| Академия » Статьи » Лекции » Анатомия |
ПРОДУКТИВНОЕ ВОСПАЛЕНИЕ 1
|
ПРОДУКТИВНОЕВОСПАЛЕНИЕ • Продуктивное, или пролиферативное, воспаление характеризуется преобладанием пролиферации клеточных элементов. Главными признаками продуктивного воспаления являются: инфильтрация мононуклеарами, особенно макрофагами, лимфоцитами и плазматическими клетками, пролиферация фибробластов, во многих случаях — нарастающий фиброз и выраженная в той или иной степени деструкция (альтерация) ткани. При этом Процессы экссудации также имеют место, но они отходят на второй план. Этиология. Причины продуктивного воспаления разнообразны — биологические факторы (вирусы, риккетсии, бактерии, грибы, паразиты), физические и химические факторы (чаще имеет место пролонгированная экспозиция неповреждающими, но потенциально токсичными субстанциями типа асбеста, оксида кремния (IV) и т.п.; инородные тела и др.)» иммунные реакции, в частности такие, которые возникают, например, против собственных тканей при аутоиммунных болезнях. Иногда непосредственно этиологический фактор вызывает продуктивную реакцию, что особенно типично для вирусов и риккетсии. Учитывая часто наблюдаемую персистенцию этиологического фактора, основными особенностями продуктивного воспаления считают длительное воздействие патогенного раздражителя и хроническое течение (только в редких случаях оно протекает остро). Длительная антигенная стимуляция, развивающаяся зачастую на фоне измененной реактивности организма, приводит к развитию реакций гиперчувствительности. Для продуктивного воспаления характерны пролиферация клеток гематогенного и гистиогенного происхождения, дифференцировка их и клеточные трансформации (схема 21). В очагах продуктивного воспаления отмечается выраженная пролиферация моноцитов. Моноциты начинают эмигрировать относительно рано и в пределах 48 ч становятся преобладающими. Достигнув экстраваскулярных тканей, моноциты трансформируются в макрофаги. Появление макрофагов осуществляется тремя механизмами. Во-первых, из циркулирующей крови. Это наиболее важный источник. Стимулом для появления моноцитов служат фибриноген, пептиды, катионные белки нейтрофилов, лимфокины, некоторые факторы роста (трансформирующий фактор роста, тромбоцитарный фактор роста), а также фрагменты разрушающегося коллагена и фибронектин. Каждый из них играет роль при определенных обстоятельствах. Например, лимфокины появляются во время иммунных реакций типа ГЗТ. Во-вторых, местная пролиферация — путем митотического деления макрофагов после эмиграции их из крови. В-третьих, пролонгированное выживание ("бессмертные клетки") и иммобилизация макрофагов в зоне воспаления. Этот вариант типичен для медленных вирусных инфекций или в случаях отложения малотоксичных веществ типа инертных липидов, угольной пыли. Макрофаг является центральной фигурой при продуктивном воспалении благодаря большому количеству биологически активных продуктов, которые он может производить. Одни из этих продуктов токсичны по отношению к тканям (например, метаболиты кислорода, протеазы), другие вызывают приток клеток иных типов (лимфоцитов, нейтрофилов, фибробластов), каждая из которых вносит свою лепту в широкую палитру изменений при хроническом воспалении, для которого характерны прогрессирующее повреждение тканей и последующие функциональные Нарушения. Макрофаг — основной представитель системы моноцитарных фагоцитов (СМФ), его нередко называют профессиональным фагоцитом, так как основной его функцией является эндоцитоз, а еще точнее, фагоцитоз. Для осуществления этой функции на поверхности макрофага имеются десятки специфических рецепторов, в частности развитая и сложная система Fc-рецепторов к различным иммуноглобулинам, лектиноподобные рецепторы для захвата бактерий, грибов и т.д. Фагоцитоз — характерная черта продуктивного воспаления, однако он далеко не всегда завершается полным перевариванием чужеродного агента. Во многих случаях живые агенты, обладая защитными механизмами, выживают внутри макрофагов (эндоцитобиоз), и процесс приобретает хронический характер. Являясь самой распространенной клеткой среди других клеток воспалительного инфильтрата, макрофаг в то же время занимает ключевую позицию в клеточных кооперациях при продуктивном воспалении (схема 22). Важнейшими продуктами секреции макрофагов являются цитокины (ИЛ-1, ИЛ-2, ИЛ-5, ИЛ-6, ИЛ-8, ФНО-а), с помощью которых осуществляются прежде всего активация лимфоцитов и межклеточные взаимоотношения, и факторы роста (ФРФ, ТцФР, КСФ, ТФР), стимулирующие клеточную пролиферацию, в частности, Т- и В-лимфоцитов, эндотелиоцитов, тромбоцитов и клеток-предшественников миелоидного ряда в костном мозге. Влияние на эндотелий заключается в том, что, с одной стороны, происходит его пролиферация, с другой — усиливается адгезия тромбоцитов. Кроме того, к секреторным продуктам макрофага относится огромное количество ферментов (кислые фосфатазы — гидролазы, липазы; эстераза, нейтральные протеазы — эластаза, коллагеназа и др.), реактивные метаболиты кислорода (пероксид водорода, супероксидный анион-радикал), биоактивные л-и -п и д ы , например производные арахидоновой кислоты (простаг-ландины, лейкотриены, ФАТ), фибронектин, цАМФ, оксид азота (II) и др. Схема 21. Пролиферация, дифференцировка, трансформация клетоквочагевоспаления 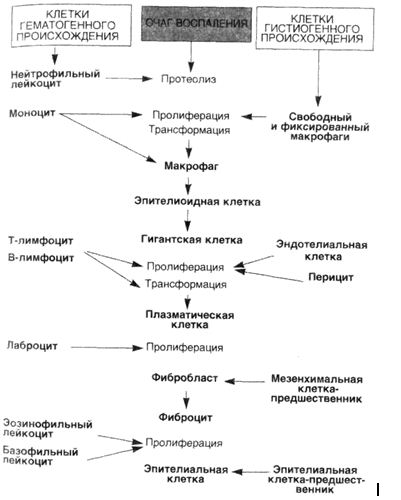 Схема 22. Взаимоотношениемакрофагасклеткамив очагевоспаления 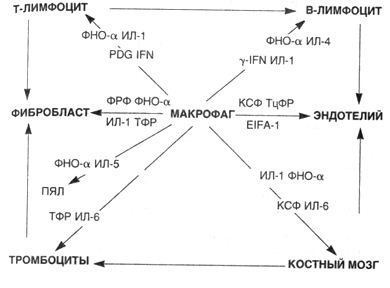 Клеточные кооперации, т.е. взаимодействие макрофага и других клеток воспалительного инфильтрата между собой, с одной стороны, и взаимодействие клеток с волокнистыми структурами и различными компонентами экстрацеллюлярного матрикса, с другой, осуществляются с помощью комплекса интегриновых рецепторов. Например, кооперация макрофаг —■ лимфоцит осуществляется с помощью интрацеллюлярных адгезивных молекул (ICAM-1). При этом в межклеточных кооперативных взаимоотношениях важную роль играют антигены гистосовместимости (система HLA): они выступают в качестве маркеров, осуществляющих антигенное считывание, а также выполняют роль универсальных рецепторов, представляющих чужеродные антигены. Другими часто встречающимися клетками, помимо макрофага, при продуктивном воспалении являются лимфоциты, плазматические клетки, эозинофилы и тучные клетки. Плазматические клетки секретируют антитела, направленные либо против персистирующих антигенов в зоне воспаления, либо против поврежденных компонентов ткани. В ряде случаев плазматические клетки превращаются в гомогенные шаровидные образования, которые называют гиалиновыми шарами, или тельцами Русселя. Лимфоциты мобилизуются как в антительных, так и в клеточных иммунных реакциях. Однако при неиммунном воспалении лимфоциты встречаются также очень часто, и причина этого явления до сих пор является загадкой для морфологов. При иммунном воспалении лимфоциты активируются, контактируя с антигенами. Активированные лимфоциты вырабатывают лимфокины — главные стимуляторы моноцитов и макрофагов, и 7-интерферон, который способствует активации и дифференцировке макрофагов. В то же время активированные макрофаги выделяют монокины (ИЛ-1), которые в свою очередь влияют на функцию Т- и В-лимфоцитов. Эозинофилы преобладают в клеточных инфильтратах прежде всего при паразитарных инфекциях, т.е. в случаях иммунных реакций, связанных с IgE. Эозинофилы относят к хемотаксическим агентам, привлекающим главным образом тучные клетки, гранулы которых содержат главный базовый протеин, очень токсичный для паразитов. Эозинофилы, хотя и являются главными при паразитарных инфекциях, вносят свой вклад в повреждение тканей при состояниях гиперчувствительности. Нейтрофильные лейкоциты обычно являются ключевым признаком экссудативного воспаления, однако в ряде случаев довольно четко "уживаются"признаки хронического продуктивного и острого (экссудативного) воспаления. Это наблюдают, например, при актиномикозе, при активном хроническом гастрите. Но наиболее значимым для течения и исхода продуктивного воспаления оказывается влияние макрофага (ФРФ, ИЛ-1, ФНО-а) вместе с тромбоцитами (ТцФР, ТФР-) и Т-лимфоцитами ФНО-а) на фибробласты. Медиаторы этих трех эффекторных клеток усиливают пролиферацию и метаболизм фибробластов и тем самым индуцируют синтез коллагена. Таким образом, еще одной существенной особенностью продуктивного воспаления является развитие склероза в исходе воспаления. Механизм, который приводит к воспроизводству и распространенной пролиферации фибробластов, к сосудистой пролиферации, накоплению коллагена и фиброзу при хроническом продуктивном воспалении, аналогичен тому, который возникает при заживлении ран (см. лекцию 16 "Воспаление, регенерация и дисрегенерация"). Выделяют следующие виды продуктивного воспаления: 1) интерстициальное (межуточное); 2) гранулематозное; 3) воспаление с образованием полипов и остроконечных кондилом. ИНТЕРСТИЦИАЛЬНОЕ (МЕЖУТОЧНОЕ) ВОСПАЛЕНИЕ Этот вид воспаления характеризуется образованием очагового или диффузного воспалительного клеточного инфильтрата в строме паренхиматозных органов — миокарда, печени, почек, легких. Инфильтрат представлен лимфоцитами, гистиоцитами, плазматическими клетками, единичными нейтрофилами, эозинофилами и тучными клетками. В паренхиматозных элементах органов выявляются выраженные дистрофические, а иногда и некробиотические изменения. Подобные изменения наблюдаются в кардиомиоцитах, например при миокардите Абрамова — Фидлера, в гепатоцитах при вирусном гепатите. В исходе хронического интерстициального воспаления разрастается соединительная ткань. При некоторых заболеваниях печени хроническое интерстициальное воспаление приводит к развитию цирроза. ГРАНУЛЕМАТОЗНОЕВОСПАЛЕНИЕ Гранулематозное воспаление — вариант продуктивного воспаления, при котором доминирующим типом клеток являются активированные макрофаги (или их производные), а основным морфологическим субстратом — гранулема. Гранулема, или узелок (бугорок, по Р.Вирхову), — это очаговое скопление способных к фагоцитозу клеток моноцитарно-макрофагальной природы. Основным представителем клеток СМФ является макрофаг, который, как уже упоминалось, образуется из моноцита. На "поле" воспаления моноцит делится лишь один раз, а затем трансформируется, как показал опыт с культурой ткани, в макрофаг. Но на этом трансформации не заканчиваются. Через 7 дней после возникновения и размножения макрофаг превращается в эпителиоидную клетку. Для этого необходимы продукты активированных Т-лимфоцитов, особенно 7-интерферон. Эпителиоидные клетки по сравнению с макрофагами имеют более низкую фагоцитарную способность (у них отсутствуют вторичные лизосомы и макрофагальные гранулы), но лучше развитую бактерицидную и секреторную активность — они синтезируют факторы роста (ФРФ, ТФР), фиброне-ктин-1, ИЛ-1. На второй неделе эпителиоидные клетки трансформируются путем деления ядер без деления клетки (реже путем слияния между собой) в гигантские многоядерные клетки Пирогова — Лангханса, а через 2—3 нед — в гигантские клетки инородных тел. Особенностями гигантских клеток Пирогова — Лангханса являются крупные размеры (до 40—50 мкм), наличие большого (до 20) количества ядер, которые располагаются эксцентрично с одной стороны в форме подковы. В гигантской клетке инородных тел ядер еще больше — до 30 (описывают даже до 100), но они располагаются преимущественно в центре клетки. Оба типа гигантских клеток отличает отсутствие лизосом, поэтому, захватывая различные патогенные факторы, гигантские клетки не в состоянии их переварить, т.е. фагоцитоз в них подменяется эндоцитобиозом. В случаях микробной инвазии эндоцитобиоз поддерживается наличием в цитоплазме секреторных гранул, напримерлипидных включений при туберкулезе. Однако в основном секреторная функция их резко подавлена, факторы роста и цитокины, в частности, вообще не синтезируются. Морфогенез гранулемы складывается из следующих четырех стадий: ▲ накопление в очаге повреждения ткани юных моноцитарных фагоцитов; ▲ созревание этих клеток в макрофаги и образование макрофагальной гранулемы; ▲ созревание и трансформация моноцитарных фагоцитов и макрофагов в эпителиоидные клетки и образование эпителиоидно-клеточной гранулемы; ▲ трансформация эпителиоидных клеток в гигантские (Пирогова — Лангханса и/или инородных тел) и формирование гигантоклеточных гранулем. Таким образом, учитывая преобладающий клеточный состав гранулемы, по морфологическим признакам различают три вида гранулем: 1) макрофагальную гранулему (простую гранулему, или фагоцитому); 2) эпителиоидно-клеточную гранулему; 3) гигантоклеточную гранулему. Этиология гранулематоза. Различают эндогенные и экзогенные этиологические факторы развития гранулем. Кэндогенным относят труднорастворимые продукты поврежденных тканей, особенно жировой ткани (мыла), а также продукты нарушенного обмена, такие как ураты. К экзогенным факторам, вызывающим образование гранулем, относят биологические (бактерии, грибы, простейшие, гельминты), органические и неорганические вещества (пыли, дымы и т.п.), в том числе лекарственные. В настоящее время гранулемы по этиологии разделяют на две группы: 1) гранулемы установленной этиологии и 2) гранулемы неустановленной этиологии [Струков А.И., Кауфман О.Я., 1989]. Первую группу в свою очередь подразделяют на две подгруппы: инфекционные и неинфекционные гранулемы. К инфекционным относят гранулемы при сыпном брюшном тифах, бешенстве, вирусном энцефалите, актиномикозе, шистосомозе, туберкулезе, лепре, сифилисе и др. Неинфекционные гранулемы развиваются при попадании в организм органической и неорганической пыли шерсть, мука, оксид кремния (IV), асбест и др., инородных тел медикаментозных воздействиях (гранулематозный гепатит, олео гранулематозная болезнь). К гранулемам неустановленной этиологии относят гранулемы при саркоидозе, болезни Крона, первичном билиарном циррозе и др. Патогенез гранулематоза. Далеко не полный перечень этиологических факторов выявляет совершенно очевидную закономерность — гранулематозное воспаление протекает, как правило, хронически и развивается при следующих двух условиях: 1 наличие веществ, способных стимулировать СМФ, созревание трансформацию макрофагов; 2) стойкость раздражителя по отношению к фагоцитам. Такой раздражитель в условиях незавершенного фагоцитоза и измененной реактивности организма оказывается сильнейшим антигенным стимулятором для макрофага и Т- и В-лимфоцитов. Активированный макрофаг с помощью ИЛ-1 еще в большей степени привлекает лимфоциты, способствуя их активации и пролиферации, — завязываются механизмы клеточно-опосредованного иммунитета, в частности механизмы ГЗТ (подробнее см. лекцию 17 "Реакции гиперчувствительности")- В этих случаях говорят об иммунной гранулеме. Иммунные гранулемы чаще построены по типу эпителиоидно-клеточных узелков, но в них всегда имеется примесь довольно большого количества лимфоцитов и плазматических клеток. Развиваются они прежде всего при таких инфекциях, как туберкулез, лепра, сифилис, склерома. Иногда продукты тканевого повреждения становятся источником антигенного раздражения и в этих случаях могут подключаться аутоиммунные механизмы гранулемообразования. Наконец, гранулемы, вызванные органическими частицам пыли и аэрозолями, содержащими белки птиц, рыб, шерсть животных, как правило, по механизму их развития являются также антигенно-опосредованными. Хотя иногда возникают механизмы образования гранулем, опосредованные антителами. К числу неиммунных гранулем относится большинство гранулем, развивающихся вокруг инородных тел, состоящих, прежде всего из частиц органической пыли (например, оксид бериллия (II) является соединением, вызывающим иммунные гранулемы саркоидного типа). Фагоцитоз в клетках неиммунных гранулем более совершенен, и построены они чаще по типу фагоцитомы либо гигантоклеточной гранулемы, состоящей из клеток инородных тел. При сравнении этих гранулем с иммунными отмечается меньшее количество лимфоцитов и плазматических клеток. К числу критериев в оценке гранулем относят показатель клеточной кинетики, т.е. степени быстроты обмена (обновления) клеток внутри гранулемы, на основании которого выделяют быстро и медленно обновляющиеся гранулемы. Быстро обновляющиеся (за 1—2 нед) гранулемы продуцируют очень токсичные вещества (микобактерии туберкулеза, лепры), построены в основном по типу эпителиоидно-клеточных, характеризуются тем, что их клетки быстро погибают и заменяются новыми, а чужеродный материал лишь частично располагается в макрофагах — все это свидетельствует об интенсивности клеточного обновления. В медленно обновляющихся гранулемах патогенный агент целиком располагается в макрофагах, при этом кинетика обмена резко замедлена. Такие гранулемы возникают при воздействии инертными малотоксичными веществами и построены чаще всего из гигантских клеток. Этот критерий важен для сравнения гранулем вокруг инородных тел экзогенного и эндогенного происхождения (шовный материал, места татуировок, неорганические пылевые частицы). Некоторые гранулемы инфекционной этиологии обладают относительной морфологической специфичностью. Для подтверждения диагноза необходима идентификация возбудителя. Специфическими называют те гранулемы, которые вызываются специфическими возбудителями (микобактерии туберкулеза, лепры, бледная трепонема и палочка склеромы), характеризуются относительно специфичными морфологическими проявлениями (только для этих возбудителей и ни для каких других), причем клеточный состав, а иногда и расположение клеток внутри гранулем (например, при туберкулезе) также довольно специфичны. |
| Вы можете прокомментировать статью | |
