| Академия » Статьи » Лекции » Анатомия |
МОРФОЛОГИЧЕСКАЯ ХАРАКТЕРИСТИКА, МОРФОГЕНЕЗ И ГИСТОГЕНЕЗ ОПУХОЛЕЙ 2
|
В большинстве органов диспластический процесс развивается при наличии пролиферации клеточных элементов на фоне предшествующейгиперплазии в связи с хроническим воспалением и дисрегенерацией. Однако в ряде случаев дисплазия сочетается с атрофией ткани, как это бывает при атрофическом гастрите с перестройкой эпителия, а также при циррозе печени. Сочетание дисплазии и атрофии не случайно, так как и тот, и другой процессы имеют общие генетические механизмы, в которых участвует ряд клеточных онкогенов, ген-супрессор р53 и др. В одних ситуациях активация данных генов приводит к апоптозу и атрофии без или в сочетании с дисплазией, в других — к пролиферации также без или в сочетании с дисплазией. На стадии дисплазии методами иммуногистохимии и молекулярной биологии регистрируются перестройки в работе онкопро-теинов, факторов роста, интегриновых рецепторов и адгезивных молекул. Причем генетические перестройки могут значительно опережать морфологические изменения и служить ранними признаками предопухолевых изменений. Стадия неинвазивной опухоли. Прогрессирова-ние дисплазии связывают с дополнительными воздействиями, ведущими к последующим генетическим перестройкам и злокачественной трансформации. В результате возникает малигнизированная клетка, которая некоторое время делится, формируя узел (клон) из себе подобных клеток, питаясь за счет диффузии питательных веществ из тканевой жидкости прилежащих нормальных тканей и не прорастая в них. На данной стадии опухолевый узел не имеет еще своих сосудов. Причина этого неизвестна. Вероятно, малая масса опухоли обусловливает недостаточную продукцию факторов, стимулирующих ангиогенез и стромообразо-вание в опухоли. Однако, по нашему мнению, представляется более верной точка зрения об отсутствии в неинвазивной опухоли определенных генных перестроек, которые необходимы для осуществления инвазивного роста. В случае рака стадия роста опухоли "самой в себе" без разрушения базальной мембраны и без образования стромы и сосудов называется стадией рака на месте — cancer in situ, и выделяется в самостоятельную морфогенетическую стадию. Длительность течения данной стадии может достигать 10 лет и более. Стадия инвазивной опухоли. Она характеризуйся появлением инфильтрирующего роста. В опухоли появляется развитая сосудистая сеть, строма, выраженная в различной степени, границы с прилежащей неопухолевой тканью отсутству-1От за счет прорастания в нее опухолевых клеток. Инвазия опухоли протекает в три фазы и обеспечивается определенными генетическими перестройками. Первая фаза инвазии опухоли характеризуется ослаблением контактов между клетками, о чем свидетельствуют уменьшение количества меж. клеточных контактов, снижение концентрации некоторых адгезивных молекул из семейства CD44 и др. и, наоборот, усиления экспрессии других, обеспечивающих мобильность опухолевых клеток и их контакт с экстрацеллюлярным матриксом. На клеточной поверхности снижается концентрация ионов кальция, что приводит к повышению отрицательного заряда опухолевых клеток. Усиливается экспрессия интегриновых рецепторов, обеспечивающих прикрепление клетки к компонентам экстрацеллюлярного матрикса — ламинину, фибронектину, коллагенам. Во второй фазе опухолевая клетка секретирует протеолитические ферменты и их активаторы, которые обеспечивают деградацию экстрацеллюлярного матрикса, освобождая тем самым опухоли путь для инвазии. В то же время продукты деградации фибронектина и ламинина являются хемоаттрактантами для опухолевых клеток, которые мигрируют в зону деградации в третьей фазе инвазии, а затем процесс повторяется снова. Стадия метастазирования. Это заключительная стадия морфогенеза опухоли, сопровождающаяся определенными гено- и фенотипическими перестройками опухоли. Процесс метастазирования связан с распространением опухолевых клеток из первичной опухоли в другие органы по лимфатическим, кровеносным сосудам, периневрально, имплантационно, что легло в основу выделения видов метастазирования. Процесс метастазирования объясняется с помощью теории метастатического каскада, в соответствии с которой опухолевая клетка претерпевает цепь (каскад) перестроек, обеспечивающих распространение в отдаленные органы. В процессе метастазирования опухолевая клетка должна обладать определенными качествами, позволяющими ей проникать в прилежащие ткани и просветы сосудов (мелких вен и лимфатических сосудов); отделяться от опухолевого пласта в ток крови (лимфы) в виде отдельных клеток или небольших групп клеток; сохранять жизнеспособность после контакта в токе крови (лимфы) со специфическими и неспецифическими факторами иммунной защиты; мигрировать в венулы (лимфатические сосуды) и прекрепляться к их эндотелию в определенных органах; осуществлять инвазию микрососудов и расти на новом месте в новом окружении. Метастатический каскад условно может быть разделен на четыре этапа: ▲ формирование метастатического опухолевого субклона; ▲ инвазия в просвет сосуда; ▲ циркуляция опухолевого эмбола в кровотоке (лимфотоке); ▲оседание на новом месте с формированием вторичной опухоли (рис. 5). Рис. 5. Метастатический каскад [Cotran R.S., Robins L.S., 1989]. БМ — базальная мембрана; ЭЦМ — экстрацеллюлярный матрикс. 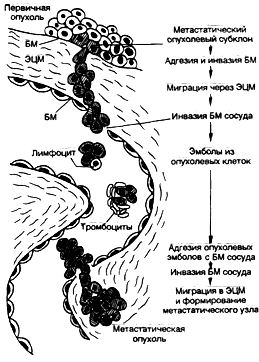 Процесс метастазирования начинается с появления метастатического субклона опухолевых клеток с измененной плазмолеммой, в результате чего клетки теряют межклеточные контакты и приобретают способность к передвижению. Затем опухолевые клетки мигрируют через экстрацеллюлярный матрикс, прикрепляясь интегриновыми рецепторами к ламинину, фибронектину, коллагеновым молекулам базальной мембраны сосуда, осуществляют ее протеолиз за счет выделения коллагеназ, катепсина, эластазы, гликозаминогидролазы, плазмина и др. Это позволяет опухолевым клеткам инвазировать базальную мембрану сосуда, прикрепляться к его эндотелию, а затем, изменяя свои адгезивные свойства (супрессия адгезивных молекул семейства САМ), отделяться как от опухолевого пласта, так и от эндотелия сосуда, "а следующем этапе формируются опухолевые эмболы, которые могут состоять только из опухолевых клеток или же из опухолевых клеток в сочетании с тромбоцитами и лимфоцитами. Фибриновое покрытие таких эмболов может защищать опухолевые клетки от элиминации клетками иммунной системы и действия неспецифических факторов защиты. На заключительном этапе опухолевые клетки взаимодействуют с эндотелием венул за счет "homing''-рецепторов и молекул семейства CD44, происходит прикрепление и протеолиз базальной мембраны, инвазия в периваскулярную ткань и рост вторичной опухоли. Клональныетеориипроисхождения иэволюцииопухоли Важнейшим в онкогенезе является вопрос о том, развивается ли злокачественная опухоль при малигнизации одной клетки или нескольких. Ответ на этот вопрос неоднозначен, так как, хотя большинство опухолей и имеет моноклоновое происхождение, существует небольшое число опухолей, формирующихся из нескольких клеток. Теория моно к л оно в о г о происхождения опухолей. У большинства опухолей такое происхождение доказывается наблюдениями неоплазм у женщин, гетерозиготных по изоформе глюкозо-6-фосфатдегидрогеназы (Г-6-ФДГ). Известно, что ген Г-6-ФДГ локализуется в Х-хромосомах, одна из которых получена от матери, а другая от отца. Одна из двух Х-хромосом, содержащихся в каждой клетке, инактивируется на стадии бластоцисты, и в клетке остается единственная Х-хромо-сома, кодирующая ту или иную изоформу Г-6-ФДГ. Это приводит к тому, что все клетки женского организма подразделяются на две группы в зависимости от изоформы Г-6-ФДГ. Описанная гетерогенность по Г-6-ФДГ свойственна многим женщинам негритянского происхождения. При этом установлено, что развивающиеся у них опухоли состоят из клеток только одного вида по изоформе Г-6-ФДГ, т.е. имеют моноклоновое происхождение. Методом инактивации Х-хромосомы доказано моноклоновое происхождение аденом и карцином толстой кишки, аденом околощитовидной железы. При хроническом миелолейкозе имеется другой маркер моноклонового происхождения опухолевых клеток — филадельфийская хромосома, при Т- и В-клеточных лимфомах и лейкозах — специфические перестройки генов Т- и В-рецепторов, выявляемые с помощью блот-анализа ДНК. Моноклоновое происхождение сопровождается ростом опухоли на начальной стадии развития в виде одного узла — т.е. имеет место уницентрической характер роста. Однако моноклоновость уже развившейся опухоли можетбыть результатом не моноклонового ее происхождения, а возникать в итоге селекции наиболее злокачественного клона опух0' левых клеток и вытеснения им менее злокачественных клонов- Теория поликлонового происхождения опухолей. Поликлоновое происхождение опухолей встречается значительно реже, чем моноклоновое, и характерно для опухолей с мультицентричным характером роста, таких как полипоз толстой кишки, мультицентрический рак молочной железы и печени и др. Мультицентрический характер роста может обусловливать и формирование единого узла опухоли при близком расположении и слиянии нескольких очагов роста. Это положение впервые обосновал Willis в своей теории опухолевого поля (1967). Согласно данной теории, возможно образование одновременно нескольких очагов пролиферации клеток с их последующей трансформацией. По мере роста опухолевые очаги сливаются. При этом опухолевые клетки принадлежат к разным клонам. В ходе прогрессии опухоли может происходить ее клональная эволюция [Nowell P., 1988], т.е. могут появляться новые клоны опухолевых клеток, возникающие в результате вторичных мутаций, что приводит к поликлоновости опухоли и доминированию наиболее агрессивных клонов как итог клональной селекции. Доброкачественные опухоли характеризуются доминированием опухолевых клеток одного клона на протяжении всего существования, в то время как в злокачественных опухолях постоянно прогрессирует поликлоновость, особенно при низкодифференцированных высокозлокачественных вариантах. Теория клональной эволюции может помочь в объяснении не только прогрессии злокачественной опухоли и метастазирования, но также дать ответы на следующие вопросы: почему в опухолях может возникать феномен "метаплазии" — изменения дифференцировки клеток на отдельных участках? Как может усиливаться злокачественность опухоли со временем или особенно после проведения противоопухолевой терапии? Почему возникают устойчивые к противоопухолевым воздействиям опухоли спонтанно и после терапевтических воздействий (феномен множественной лекарственной устойчивости опухоли). Взаимодействиеопухолииорганизма-опухоленосителя Взаимодействие опухоли и организма-опухоленосителя складывается из действия опухоли на организм и защиты организма-опухоленосителя от опухоли. Действие опухоли на организм хозяина. Проявляется в локальном и общем воздействии. Локальное воздействие включает в себя нарушения метаболизма, сдавление прилежащих и разрушение растущей опухолью сохранных тканей, прорастание стенок сосудов, что может приводить к местному венозному застою. Некроз и изъязвление опухоли могут сопровождаться кровотечением, присоединением вторичных инфекций. Общее воздействие опухоли на организм хозяина может приводить к развитию различных анемий, раковой интоксикации, раковой кахексии и паранеопластических синдромов. Остановимся на двух последних проявлениях. Раковая кахексия. Характеризуется истощением больного с развитием бурой атрофии миокарда, печени и скелетной мускулатуры. Возникновение раковой кахексии связывают с увеличением уровня белкового обмена в ткани опухоли, которая становится ловушкой всех питательных веществ и обрекает организм на "голодание". В последние годы развитие раковой кахексии связывают также с усиленной продукцией α-ФНО макрофагами и другими клетками в организме-опухоленосителе. а-ФНО называется кахектином, и его патогенетическая роль в возникновении кахексии доказана пока только в экспериментах на животных. Паранеопластические синдромы. Это синдромы, обусловленные наличием опухоли в организме. Патогенез их различен, но всегда ключевое событие обусловлено воздействием неоплазмы. При гормонально-активных опухолях могут возникнуть различные эндокринопатии, как, например, синдром Иценко — Кушинга при аденомах передней доли гипофиза или нейроэндокринных опухолях легких; гиперкальциемия и остеопороз при аденомах околощитовидных желез и раке легкого. Опухоль, особенно на стадии метастазирования, воздействует на свертывающую и противосвертывающую систему крови, сама продуцирует факторы, усиливающие коагуляцию крови, и способствует развитию различных тромбопатий (мигрирующие тромбофлебиты, небактериальный тромбоэндокардит) и афибриногенемии. При опухолевом росте описан широкий спектр иммунопатологических процессов с иммунокомплексными, анти-тельными и цитотоксическими механизмами развития. Эти процессы приводят к возникновению у онкологических больных нейропатий, миопатий и дерматопатий. Механизмы противоопухолевой защиты организма. Они разнообразны и складываются из защитных реакций с участием процессов репарации мутированного участка ДНК, сбалансирован' ной работы генов-супрессоров и клеточных онкогенов, а также из факторов неспецифической и специфической, иммунной и неиммунной защиты. 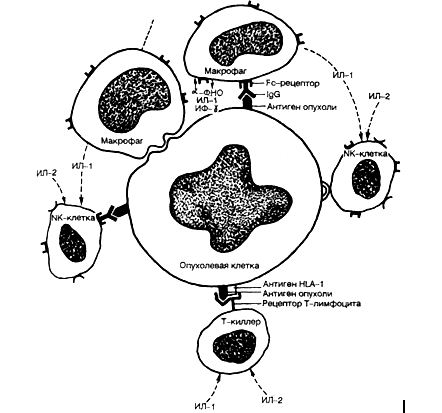 Рис. 6. Противоопухолевая иммунная реакция. В защите от опухоли имеют значение реакции клеточного и гуморального иммунитета (рис. 6). Основными клетками, участвующими в противоопухолевой иммунной защите, являются специфические цитотоксические Т-лимфоциты, способные распознать мембраносвязанные опухолевые антигены (см. лекцию 20), а также антигены, относящиеся к антигенам I класса основного комплекса гистосовместимости; натуральные Т-киллеры (NK-клетки), вызывающие деструкцию опухолевых клеток предварительной сенсибилизации. Лизис осуществляется за счет непосредственного связывания с опухолевыми клетками или через Fc-фрагменты противоопухолевых антител; макрофаги осуществляют как неспецифическое повреждение опухолевых клеток через выделение а-ФНО и других факторов, так и специфическое иммунное повреждение путем присоединения к Fc. фрагментам противоопухолевых антител и через активацию Т-лимфоцитов, выделяющих 7-интерферон и другие цитокины. Антительный механизм противоопухолевого иммунитета может осуществляться связыванием противоопухолевыми антителами комплемента с формированием литического комплекса, который взаимодействует с опухолевой клеткой, присоединением к опухолевой клетке через Fc-фрагмент противоопухолевых антител NK-клеток и макрофагов. Интересным является вопрос о неэффективности иммунных реакций в защите от опухоли. Эту неэффективность, вероятно, можно объяснить развитием иммуносупрессии у онкологических больных, наличием феномена антигенного ускользания опухоли в связи с антигенной лабильностью, антигенным упрощением ее клеток, а также усилением роста опухоли под влиянием противоопухолевых антител. ГИСТОГЕНЕЗОПУХОЛЕЙ Термин "гистогенез опухолей" означает тканевое происхождение опухоли, что не совсем точно, так как в настоящее время мы можем нередко определять не только тканевое, но и клеточное происхождение неоплазмы, т.е. ее цитогенез. Особенно хорошо изучен цитогенез опухолей кроветворной и лимфоидной ткани — гемобластозов. В основе теории цитогенеза гемобластозов положено учение о стволовых и полустволовых клетках-предшественницах кроветворения. Многие вопросы происхождения солидных опухолей остаются пока еще спорными, поскольку имеется мало данных о клетках-предшественницах многих тканей. Предлагаются гипотетические схемы цитогенеза рака легкого, желудка, молочной железы и др. В теории гисто- и цитогенеза опухолей следует выделить несколько основных аспектов. 1. Трансформации может подвергаться только пролиферирующая соматическая клетка, т.е. поли- или унипотентные клетки-предшественницы. 2. Опухолевая клетка способна повторять в извращенной форме признаки дифференцировки (т.е. фенотип), заложенные в клетке-предшественнице, из которой она возникла. 3. В опухолевых клетках извращение дифференцировки связано с наличием блока дифференцировки. При наличии такого блока дифференцировки на уровне унипотентных клеток-предшестественниц опухолевые клетки обладают минимальной фенотипической гетерогенностью. При наличии блока дифференцировки на уровне полипотентных клеток-предшественниц в неоплазме выражена фенотипическая гетерогенность опухолевых клеток и появляются клетки-химеры с множественной дифференцировкой. 4. Дифференцировка опухолевых клеток зависит как от уровня малигнизации клетки-предшественницы, так и от уровня блока дифференцировки. Доброкачественные опухоли развиваются при трансформации унипотентных клеток-предшественниц с низким блоком дифференцировки, поэтому они построены из зрелых клеточных элементов. Злокачественные опухоли характеризуются меньшим уровнем дифференцировки их клеток по сравнению с доброкачественными, что связывается с их развитием из полипотентных клеток-предшественниц и наличием высокого блока дифференцировки. Чем выше уровень малигнизации и уровень блока дифференцировки, тем менее дифференцированна возникающая злокачественная опухоль. В качестве гисто- и цитогенетических маркеров опухолевых клеток могут использоваться ультраструктурная организация опухолевой клетки, а также генные, хромосомные, антигенные и биомолекулярные маркеры, получившие название "опухолевые маркеры". В группу биомолекулярных опухолевых маркеров относят различные молекулы (факторы роста, рецепторы, онкобелки, адгезивные молекулы, интегриновые рецепторы), ферменты, белки промежуточных филаментов, рецепторов и адгезивных молекул. |
| Вы можете прокомментировать статью | |
